
Меню сайта
Наши новости
Распространение алкалоидов в растительном мире.
Умягчение воды
Изучение роста и биосинтеза бас полученными штаммамиБиблиотека / Разработка методов биотехнологического получения белков, аминокислот и нуклеозидов, меченных стабильными изотопами 2Н и 13С с высокими степенями изотопного обогащения / Библиотека / Разработка методов биотехнологического получения белков, аминокислот и нуклеозидов, меченных стабильными изотопами 2Н и 13С с высокими степенями изотопного обогащения / Изучение роста и биосинтеза бас полученными штаммами Изучение роста и биосинтеза бас полученными штаммамиСтраница 5
Изучение ростовых и биосинтетических характеристик В.
subtilis
на средах,
содержащих тяжёлую воду и гидролизаты метилотрофных бактерий.
Кривые, отражающие динамику роста, ассимиляции глюкозы и накопление инозина в культуральной жидкости штаммом В. subtilis в условиях протонированной среды и среды, с 99,9 ат.% тяжёлой воды представлены на рис. 2.
Как видно из рис. 2, при переносе клеток со стандартной на дейтерированную среду выход микробной биомассы, продолжительность лаг-фазы и длительность времени клеточной генерации в целом изменяются незначительно. При росте исходного штамма В. subtilis па среде, содержащей обычную воду уровень накопления инозина в культуральной жидкости достигал величины 17,3 г/л после пяти суток культивирования (рис. 2). Уровень накопления инозина на дейтерированной среде был снижен в 4,4 раза по-сравнению с исходным штаммом на протонированной среде (рис. 2). Низкие уровни секреции инозина на дейтерированной среде коррелируют со степенью конверсии глюкозы в этих условиях. Так, кривая конверсии глюкозы на полностью дейтерированной среде имела меньший угол наклона, чем на среде с обычной водой, что свидетельствует о том, что при росте на дейтерированной среде скорость ассимилиляции глюкозы несколько замедляется (рис. 2).
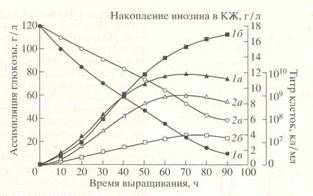
Рис.2. Динамики роста B.subtilis (1a, 2a), конверсии глюкозы (1б,2б) и накопления инозина в культуральной жидкости (1в,2в) на средах с различным изотопным составом: 1 а,б,в-B. Subtilis на обычной протонированной среде; 2 а,б,в-B.subtilis на полностью дейтерированной среде с гидролизатом дейтеро-биомассы метилотрофных бактерий.
Полученные для исследуемых микроорганизмов данные, в целом, подтверждают устойчивое представление о том, что адаптация клетки к тяжёлой воде является фенотипическим явлением, поскольку адаптированные к тяжёлой воде клетки возвращаются к нормальному росту и биосинтезу в протонированных средах после некоторого лаг-периода. В то же время обратимость роста на D2O/H2O-cpeдax теоретически не исключает возможности того, что этот признак стабильно сохраняется при росте в тяжёлой воде, но маскируется при переносе клеток на дейтерированную среду. Можно предположить, что клетка реализует лабильные адаптивные механизмы, которые способствуют функциональной реорганизации работы ферментных систем в тяжёлой воде. Также не исключено, что наблюдаемые при адаптации эффекты связаны с образованием в тяжёлой воде более прочных и стабильных связей, чем связей с участием водорода. По теории абсолютных скоростей разрыв С-Н-связей может происходить быстрее, чем C-D-связей, подвижность дейтерия D
+
меньше, чем подвижность протия Н+, константа ионизации D2O в 5 раз меньше константы ионизации Н2О (Crespy J., Kalz H.H., 1979). С точки зрения физиологии, наиболее чувствительными к замене протия на дейтерий могут оказаться аппарат биосинтеза макромолекул и дыхательная цепь, т. е. именно те клеточные системы, которые используют высокую подвижность протонов и высокую скорость разрыва водородных связей.
Смотрите также
Применение органических реагентов в аналитической химии
Органические
вещества широко применяются в аналитической химии вообще и в фармацевтическом
анализе, в частности. Ещё с начала нашей эры было известно, что настой
чернильных дубильных орешко ...
Нефть и продукты её переработки
...
Получение медноаммиачного волокна (целлюлозы) химическим методом
Среди
различных видов искусственного волокна, которые изготовляются из целлюлозы,
медноаммиачное волокно занимает особое место. Этот вид искусственного волокна
впервые был по лучен давно: е ...