
Меню сайта
Наши новости
Распространение алкалоидов в растительном мире.
Умягчение воды
Белки. Строение и общие
свойства аминокислотДипломы, курсовые и прочее / Белки и нуклеиновые кислоты / Дипломы, курсовые и прочее / Белки и нуклеиновые кислоты / Белки. Строение и общие
свойства аминокислот Белки. Строение и общие
свойства аминокислотСтраница 1
Основной структурной единицей белков являются a-аминокислоты. В природе встречается примерно 300 аминокислот. В составе белков найдено 20 различных a-аминокислот (одна из них – пролин, является не амино-, а иминокислотой). Все другие аминокислоты существуют в свободном состоянии или в составе коротких пептидов, или комплексов с другими органическими веществами.
a-Аминокислоты представляют собой производные карбоновых кислот, у которых один водородный атом, у a-углеродного атома замещен на аминогруппу (–NН2), например:

Различаются аминокислоты по строению и свойствам радикала R. Радикал может представлять остатки жирных кислот, ароматические кольца, гетероциклы. Благодаря этому каждая аминокислота наделена специфическими свойствами, определяющими химические, физические свойства и физиологические функции белков в организме.
Именно благодаря радикалам аминокислот, белки обладают рядом уникальных функций, не свойственных другим биополимером, и обладают химической индивидуальностью.
Значительно реже в живых организмах встречаются аминокислоты с b- или g-положением аминогруппы, например:
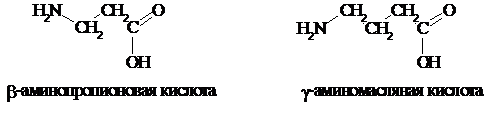
В молекулах всех природных аминокислот ( за исключением глицина) у a-углеродного атома все четыре валентные связи заняты различными заместителями, такой атом углерода является асимметрическим, и получил название хирального атома. Вследствие этого растворы аминокислот обладают оптической активностью – вращают плоскость плоскополяризованного света. Причем, при прохождении через них поляризованного луча происходит поворот плоскости поляризации либо в право (+), либо влево (–). По расположению атомов и атомных группировок в пространстве относительно асимметрического атома различают L- и D-стереоизомеры аминокислот. Знак и величина оптического вращения зависят от природы боковой цепи аминокислот (R-группы).
Число возможных стереоизомеров ровно 2n, где n – число асимметрических атомов углерода. У глицина n = 0, у треонина n = 2. Все остальные 17 белковых аминокислот содержат по одному асимметрическому атому углерода, они могут существовать в виде двух оптических изомеров.
В качестве стандарта при определении L и D-конфигураций аминокислот используется конфигурация стереоизомеров глицеринового альдегида.
Расположение в проекционной формуле Фишера NH2-группы слева соответствуют L-конфигурации, а справа – D-конфигурации.

Следует отметить, что буквы L и D означают принадлежность того или иного вещества по своей стереохимической конфигурации к L или D ряду, независимо от направленности вращения.
В составе белков обнаруживаются только L-изомеры аминокислот. D-формы аминокислот в природе встречаются редко и обнаружены лишь в составе белков клеточной стенки (гликопротеинов) некоторых бактерий и в пептидных антибиотиках (грамицидин, актиномицин и т.д.). L-формы хорошо усваиваются растениями и животными и легко включаются в обменные процессы. D-формы не ассимилируются этими организмами, а иногда даже ингибируют процессы обмена. Это объясняется тем, что ферментативные системы организмов специфически приспособлены к L формам аминокислот.
L и D формы аминокислот оказывают различное физиологическое воздействие на организм человека – различаются по вкусу: D-изомеры сладкие, L-формы горькие или безвкусные.
Взаимопревращение D и L-энатиомеров называется рацемизацией. Превращение D Û L – это один из метаболических процессов в живых организмах, причем равновесие этого метаболического процесса сильно смещено в сторону образования L-формы. Когда метаболические процессы после смерти организма прекращаются, процесс D Û L продолжается самопроизвольно с очень малой скоростью, переводя для каждой аминокислоты к соотношению D/L-энантиомеров, характерному для неметаболического равновесия. Для достижения такого равновесия могут потребоваться десятки тысяч лет. Новый метод определения геологического возраста образца основан на измерении соотношения D/L-энантиомеров аспарагиновой кислоты в образцах окаменелых костей. Результаты, полученные методом D /L-датирования, хорошо дополняют другие данные, полученные, например, радиоуглеродным методом.
Кроме 20 стандартных аминокислот встречающихся почти во всех белках, существуют еще нестандартные аминокислоты, являющиеся компонентами лишь некоторых типов белков – эти аминокислоты называют еще модифицированными. Около 150 из них уже выделены. Эти аминокислоты образуются после завершения синтеза белка в рибосоме клеток путем посттрансляционной химической модификации.
Страницы: 1 2
Смотрите также
Химическая физика и некоторые проблемы биологии
...
Исследования химии в 20-21 веках
На рубеже тысячелетий в каждой из главных
областей естествознания – биологии, физике, химии - произошли и происходят
одинаково важные, капитальные, но притом различные метаморфозы.
Бурно ...